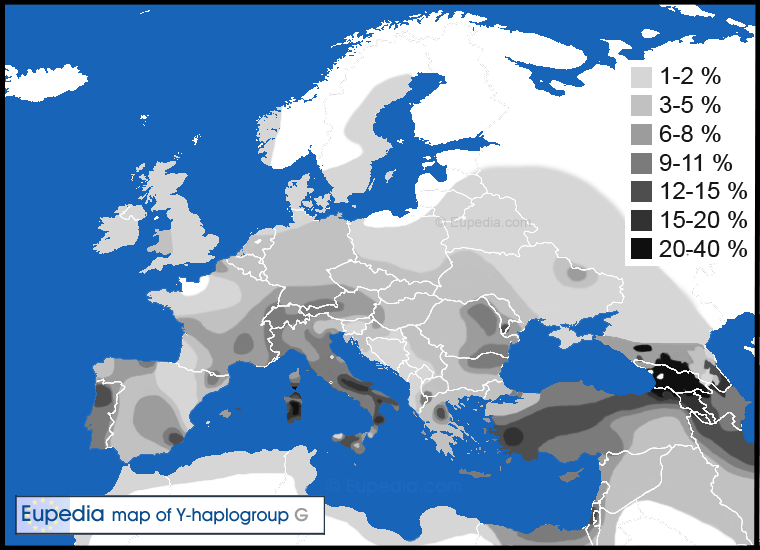
現在單倍群G儘管只有低分布頻率,但一路從西歐和北非到中亞、印度與東非都被發現(通常都佔人口中1到10%之間)。唯一的例外在高加索地區、南義大利與薩丁尼亞,這些地方的父系血脈分布頻率一貫在15%到30%之間。
壓倒性地多數歐洲人屬於G2a子分支,而多數北歐與西歐人更特別屬於G2a-L140(或是下游擴充的G2a-M406)。幾乎所有G2b(L72+,之前的G2c)都在歐洲的中歐猶太人中被發現。G2b被發現來自中東到巴基斯坦,並且幾乎都被確定來自有已經被檢定至少9250歲的G2b樣本(Broushaki et al.在2016年)的伊朗西部新石器時代農夫。
單倍群G1最主要在伊朗被發現,但也能在黎凡特、中歐猶太人中、以及中亞(尤其在哈薩克)被發現。
G2a組成了歐洲地中海沿岸5到10%的人口,但在北歐則相對稀有。歐洲唯一單倍群G2超過10%人口的地區在西班牙北部的坎塔布里亞,在葡萄牙,在義大利中部與南部(尤其在亞平寧山脈),在薩丁尼亞,在希臘北部(色薩利),在克里特島,在加告茲人當中:都在山多與相對孤立的地區。其他分布頻率接近10%的地區包括西班牙北部的阿斯圖利亞斯、法國中部的奧弗涅、瑞士、愛琴海群島與塞浦路斯。
單倍群G2a分布圖
單倍群G2a系統發生圖
單倍群G是被認為是自智人來開非洲以來在60000年前左右第二龐大的移民,巨集單倍群F的後代,儘管更早之前單倍群C與D的移民沿著南亞沿海前往更遙遠的大洋洲與遠東,單倍群F穿過了阿拉伯半島並定居在中東。他主要的分支巨集單倍群IJK變成歐亞大陸80%人口的祖先。單倍群G約在50000年前形成單倍群IJK的旁系血脈,但似乎起初發展緩慢,10000年都在孤立中發展,可能在近東被對歐亞大陸的殖民浪潮隔絕。
有303個突變(核甘酸多型性)可以定義單倍群G,確定這個父系血脈在分出單倍群G1與G2前經歷了嚴重的瓶頸。G1在26000年前左右的末次冰盛期開頭就起源於現代伊朗。G2也同時的西亞周遭發展。那時候的人類都是獵人兼採集者,而且多數案例都生活在游牧或半游牧部落。單倍群G2的成員出現剛好聯繫到早期農業在11500年前新月沃土的發展。G2a分支擴張到安那托利亞、高加索地區與歐洲,同時G2b正從伊朗橫跨新月沃土到巴基斯坦擴散。在現在最容易在黎巴嫩人與猶太人當中發現,但也低頻率分布在阿拉伯半島、敘利亞、伊拉克、伊朗、阿富汗與巴基斯坦。
至今為止被做過古人Y染色體分析,有早期新石器時代安那托利亞、伊朗、以色列、約旦以及大多數歐洲新石器時代文化(希臘的新石器時代色薩利文化、匈牙利/克羅埃西亞的斯塔爾切沃文化、德國的線性陶器文化、義大利的雷梅德洛還有法國西南部與西班牙的鳥蛤陶器文化),而且除了在黎凡特外,所有遺跡都屬於G2a個體佔多數。這強力意味著農業至少是從安那托利亞/伊朗由單倍群G成員傳播到歐洲的。被測試過的44個近東樣本中,包括來自約旦與伊朗西部的新石器時代農夫(Lazaridis et al.在2016年),被發現一位來自前陶新石器時代的G2b樣本(大約在西元前7250年)以及一位來自陶器新石器時代早期的G2a1樣本(大約在西元前5700年),都來自伊朗。少數來自黎凡特的樣本屬於單倍群CT、E1b、H2與T,但依舊無法排除單倍群G可能出現在其他樣本。13位來自安那托利亞西北部的Barcın遺址的新石器時代早期農夫的測試中(Mathieson et al.在2015年),八位屬於單倍群G2a(子分支G2a2a-PF3146、G2a2a1b-L91、G2a2a1b1-PF3247、G2a2b-L30、G2a2b2a-P303、G2a2b2a1c-CTS342)。其他樣本屬於C1a2、H2、I、I2c與J2a。這些少數單倍群也展示了在巴爾幹半島的新石器時代早期農夫之間又屬G2a占多數。偶爾會出現其他近東血脈,比如線性陶器文化的一個T1a,西班牙西北部的一個R1b-V88。T1a部落被認為在札格羅斯山脈裡馴養了羊類,同時R1b部落也在新月沃土北方馴養了家牛。
單倍群G被發現基因最多樣性在新月沃土的北部,黎凡特與高加索地區之間,很好地指示出其起源地區。被推斷新石器時代早期農夫從美索不達米亞北部擴張,西至安那托利亞與歐洲,東抵南亞,南達阿拉伯半島與非洲東北部。至今只有不屬於子分支G-P15與G-L149。1下游的G2a人群,只能被發現在南高加索地區。
新石器時代農夫與山中放牧人
在歐洲各地新石器時代遺體的檢測確認了單倍群G2a曾是9000到6000年間自安那托利亞移入歐洲的新石器時代農夫與放牧人的佔優血脈。
穀類與豆類農業的第一次發展在約11500年前的新月沃土,在現在的以色列/巴勒斯坦、約旦、黎巴嫩、敘利亞與伊拉克,但是在頭兩千五百年內沒有擴張跨出這片區域太多。推遲的原因是對允許自力更生而言早期農業太過於原始並且僅僅只能作為獵人兼採集者的其中一種飲食補給來源。種植技術藉由小麥、無花果與豆類展開。馴化小麥與大麥需要漫長過程尋求具有更大,不易脆與不會碎散的穀穗突變。美索不達米亞的沖積平原對原始穀物農業來說是理想的,因為他們不需要灌溉。
陶器第一次出現在近東大約在9000年前左右的美索不達米亞北部。陶器的發展似乎與G2a農夫向安那托利亞與歐洲的突然擴張吻合。陶器可以更好儲存穀物與豆類並且能促進與比鄰的羊類與家牛放牧人還有養豬戶之間交易。一些山羊與綿羊在11000年前,在新月沃土北緣的札格羅斯山與托魯斯山中被第一次馴養,但是到8500年前也就是陶器出現後為止都沒有被推廣到黎凡特。
在安那托利亞中南部的新石器時代聚居點加泰土丘由也帶入馴化山羊與綿羊的穀物與莢果農夫建立。僅幾世紀後(大約西元前6500年)家牛也被推廣到加泰土丘與安那托利亞中部的其他地方,想必是跟他們東方的鄰居交易來的。同樣在8500年前左右,依據對古人的基因組序測,G2a新石器時代農夫抵達了安那托利亞西北部與希臘中部的色薩利(Mathieson et al.與Hofmanova et al.在2015年)。來自新石器時代色薩利文化的G2a農夫快速擴張到巴爾幹半島與多瑙河流域。在西元前5800年抵達塞爾維亞、匈牙利與羅馬尼亞,在西元前5500年抵達德國,並且在5200年前抵達比利時與法國北部。來自位於匈牙利與克羅埃西亞的斯塔爾切沃文化(西元前6000到4500年),與來自位於匈牙利與德國的線性陶器文化的古人骸骨,都被確定G2a仍是主要的父系血脈(都是G2a2a與G2a2b),即便經過了農夫與當地人口的交融。
7800年前左右,製作鳥蛤陶器的農夫帶著家羊與家豬抵達安那托利亞西北部的瑪爾馬拉海。這群人乘船橫跨了愛琴海並且殖民了義大利半島、伊利里亞海岸、法國南部與伊比利亞半島,建立了鳥蛤陶器文化(西元前5000到1500年)。鳥蛤陶器文化的古人DNA再次測得G2a佔多數,G2a的分布頻率超過80%(相對在中歐與南歐東部有50%)。
儘管如此,數量龐大的其他少數單倍群緊接在G2a後建立了不同的新石器時代遺址,包括C1a2、H2、I*、I2a1、I2c與J2a在安那托利亞,C1a2、E-M78、H2、I*、I1、I2a、I2a1、J2、T1a在南歐東部與中歐(斯塔爾切沃文化、Sopot文化、線型陶器文化),以及E-V13、H2、I2a1、I2a2a1與R1b-V88在西歐(鳥蛤陶器文化、巨石文化)。H2與T1a在前陶新石器時代黎凡特被發現並不可否認地與G2a一同被聯繫到早期農業的發展。C1a2也在西班牙巨石文化中被發現(Olalde et al.在2014年),而且做為與第一批舊石器時代歐洲人中最老的血脈,也可能在新石器時代前的全歐洲與安那托利亞被發現。E1b1b也在前陶新石器時代黎凡特被發現,不過子分支所屬可能並非E-M78或E-V13(更可能是E1b1b1*或E-M123)。R1b-V88也確定從近東傳播,雖然是經過不同路徑,帶著家牛放牧人經過北非,然後跨過伊比利亞半島。其餘部分代表來自安那托利亞西部(I*、I2c、J2)與歐洲(E-V13、I*、I1、I2a、I2a1、I2a2)的巨石文化後代被同化後的獵人兼採集者。有趣的地方在許多這些血脈,比如C1a2、H2與I*現在都看似在任何地方絕跡了,而且部份其他的在歐洲變得非常稀有(I2c、R1b-V88)。
農業從中東到歐洲的擴張
被測試過的幾位新石器時代與銅石並用時代的巴爾幹人個體中(Mathieson et al.在2017年)發現了支系群G-L91(在新石器時代中期保加利亞)、G-P303(在新時代中期保加利亞與特里波里文化)、G-Z1903(在銅石並用時代保加利亞)、G-L42(在特里波里文化)與G-PF3359(在瓦爾納文化)。
冰人奧茨,在銅石並用時代死於5000年前左右的義大利阿爾卑斯山,歐洲保存最好的天然木乃伊,屬於單倍群G2a2a2(L91),被發現是在今天的中東,南歐(尤其是西西里島、薩丁尼亞與科西嘉)與北非相對稀有的子分支,G2a2(G-PF3146)也果不其然地被發現從黎凡特到西歐都維持低分布頻率。綜上所述,歐洲新石器時代農夫勢必屬於G2a2a(G-PF3146)與G2a2b(G-L30)與他們的子分支。
如今G2a最常在歐洲的多山地區發現,比如義大利的亞平寧山脈(15到25%)與薩丁尼亞(12%)、西班牙北部的坎塔布里亞(10%)與阿斯圖里亞斯(8%)、法國東南部的奧弗涅(8%)與普羅旺斯(7%)、瑞士(7.5%)、波希米亞山區(5到10%)、羅馬尼亞(6.5%)與希臘(6.5%)。南歐的丘陵地形在新石器時代早期G2a人們帶來山羊時確實變得更理想地適合放牧。不過最有可能解釋就是在銅石並用時代晚期與青銅器時代古印歐語系使用者從俄羅斯與烏克蘭的草原入侵歐洲後,山地為G2a部落提供了庇護。
草原人群幾乎是家牛與馬匹放牧人,並且一開始只會定居在平坦地區如匈牙利平原、歐洲大平原北部與波羅的海三國。甚至之後抵達西歐,他們依舊相對喜歡坡度低的地區比如低地三國、法國西部與不列顛群島,這些地方的R1b血脈現在都超過60%,而且有些地方還佔了80%人口。事實上,G2a百分比率最高的地方都被發現是最晚被R1a與R1b人群佔據的地方。印歐語族直到西元前1800年都沒有進入伊比利亞半島並且直到西元前1200年都沒覆蓋過整個半島,小群的G2a生存於格外封閉的地區如庇里牛斯山脈、坎塔布里亞山脈或拉曼恰的乾燥高原。古意大利人在西元前1300年前才跨越阿爾卑斯山進入義大利並且更密集地住在北部,解釋了R1b在現代義大利的南北差距,在這裡分布幾乎與新石器時代的單倍群如G2a與T1a相反。不說印歐語系語言的薩丁尼亞人則要等至羅馬人征服,只在兩千年內。
G2a所有子分支的分布地圖不僅訴說古印歐語系族群如何徹底地在歐洲的北半部淘汰G2a血脈,因為古印歐語系族群也帶著一個已經在黑海草原一帶被同化的一類G2a。這支被印歐化的G2a血脈在大舉移民前屬於G2a-L140的下游分支群如G-L13與G-Z1816(見後列表)。如今G2a的新石器時代分支群尤其被發現在安那托利亞、巴爾幹半島南部、亞平寧山脈、法國中部與庇里牛斯山脈。唯一以小比例出現在歐洲北半部的G2a,以印歐語系化的G2a分支群為主。
G2a人群可能是所有人類當中第一批得到淺色皮膚等位基因的人。對來自7000年前西班牙北部的獵人兼採集者的測試依舊有深色皮膚。對比來自巴爾幹半島與德國的新石器時代早期農夫已經擁有在現代歐洲人身上找到的淺色皮膚的等位基因。目前還不清楚淺色皮膚出現在確切哪個時間與單倍群身上,不過這表明來自穀物農業帶來的新飲食導致了維他命D的不足,以往在獵人兼採集者之間一向從魚類與肉類身上攝取。選擇更淺的膚色突變讓新石器時代農夫們為了補償對肉類的缺乏能夠透過陽光刺激維他命D生產。
G2a的印歐語系分支
與其他更偏向廣布山區的G2a分支相反,一些G2a-L140子分支被發現始終平均分布歐洲甚至新石器時代農夫只帶來極小影響的斯堪地那維亞與俄羅斯。更重要的,G2a-L140與他的子分支一樣在高加索,中亞與印度各處,尤其在來自青銅器時代印歐語系入侵者的更高種姓階級者之間。G2a-L140在橫跨歐洲到印度的出現成為了印歐語系擴散的強力論據。不過G-L140自己只出現了11000年左右的前陶新石器時代過於古老到不可能屬於印歐語系人群。只能確認更下游的子分支才可能進入黑海-裏海草原然後在顏那亞時代被草原放牧人吸收,並且跟著印歐語族移民被重新分散到整個歐洲跟亞州。這裡必須羅列出每個在青銅器時代早期擴張並且散布到北歐,中亞與南亞的子分支。當中最主要的是:
單倍群G2a-L140系統發生圖
G-L1264,被發現在北高加索、波羅的海三國、斯拉夫人與日耳曼人國家以及中亞與印度。8000年前形成,不過最近共同祖先只有4500歲,他與單倍群R1a一同傳播(古斯拉夫-波羅的語族與古印度-伊朗語族)。
單倍群G-U1分布圖
G-L13存在了10500年左右,不過所有存活的攜帶者都來自生活在最多5000年前的最近共同祖先,對應到顏那亞時代。儘管年紀尚輕,他還是在整個歐洲包括俄國、還有中亞、伊朗、高加索地區語黎凡特被發現。這個分支與單倍群R1a與R1b共同被傳播。
單倍群G-L497分布圖
G-Z1816,在整個西歐與中歐被發現,尤其是日耳曼國家。他的總結年紀被Yfull估計在4500年左右,不過他的子分支G-L42在生存於特里波里文化外圍一個4900到5600歲左右的個體中被發現。這個支系也許在古印歐語系人群接觸到移居黑海草原並與顏那亞部落融合的特里波里文化晚期人群時被同化的。並且在稍後與R1b一起傳播到日耳曼與塞爾特國家。
R1b1a1a2(M269)與前古印歐語系使用者的發源地被推定定位在安那托利亞東部與/或北高加索地區。高加索地區本身也是單倍群G的熱點。所以完全可以想像屬於單倍群G(然後可能有J2b)的高加索的少數族群融合進橫跨高加索山脈並且於西元前7000到4500年之間建立在黑海北岸與東岸的R1b社群。
另一個理論認為G2a-L140來自新石器時代的安那托利亞與東歐中部(DNA檢測證明的事實)。當屬於G-U1分支的南歐東部人在現代的摩爾多瓦周遭建立庫庫泰尼-特里波里文化(包括其他人的單倍群尤其I2a1b-L621)。庫庫泰尼-特里波里人群與鄰近的草原文化積極地進行交易,然後從西元前3500年起,在顏那亞時代的黑海-裏海草原,庫庫泰尼-特里波里人群開始向東擴張到現在烏克蘭西部,離開他們的城鎮(在當時是全世界最大的),並且選擇越來越偏向他們的顏那亞鄰居一樣的游牧生活。到古印歐語系人群開始大量擴張,包括G-L13與G-L1264子分支在內的G2a-U1人群在R1b與R1a部落入侵歐洲,還有中亞與南亞時加入了他們。
G2a的庫拉-阿拉斯分支
印歐語系族群的起源同時在黑海草原形成於邁科普文化(西元前3700到3000年)與顏那亞文化(西元前3500到2300年),另一個青銅器時代社會在高加索山脈開始發展:庫拉-阿拉斯文化(西元前3400到2000年)。雖然庫拉-阿拉斯人群更少有軍事活動且更慣於定棲,也經歷過一次大擴張,首先西至安那托利亞,南抵新月沃土並且東達可能通往巴基斯坦的伊朗平原,並且可能影響了印度河文明。庫拉-阿拉斯文化的後代可能殖民了希臘島嶼,包括在克里特島建立了歐洲最古老的米諾斯文明(西元前2600到1100年),米諾斯文明遺體包括G2a-P303>pre-L140(PF3337+,G-L140)與J2a1a-L26>J-Y7010>J-Y13128>J-Z36834。
單倍群G-L293分布圖
根據現代的系統發生圖,庫拉-阿拉斯人群被認為主要屬於Y染色體單被群J2a1(最大的血脈)、J1a2-Z1828、T1a-P77、G2a1(G-L293,或稱G-FGC7535或G-Z6552)、G2a2b1a(G-M406)與L1b。其他少數單倍群也有出現,包括現代稀有的R1b1a-L388,確定屬於一位庫拉-阿拉斯個體(Lazaridis et al.在2016年)。在古典時代的希臘島嶼居民就是米諾斯人的多數後代,殖民南義大利並且帶入他們的庫拉-阿拉斯血脈給當地。
最古老的G-L293樣本來自新石器時代的西部伊朗。如今,G-L293是北高加索最普遍的G2a分支群,在北奧塞梯佔高達64%人口。庫拉-阿拉斯文化從G-L293相當稀少的南高加索地區擴張(1%在亞美尼亞與土耳其東部)。儘管如此還是能在土耳其、伊朗西北部、伊拉克北部、敘利亞與黎巴嫩發現其低分布頻率(換句話說這些地方都曾屬於庫拉-阿拉斯文化範圍),甚至也在保加利亞、羅馬尼亞、希臘、南義大利(尤其西西里島)與科西嘉。G-L293明顯沒出現在薩丁尼亞與義大利北半部,始之排除了與新石器時代和印歐語系的起源。
單倍群G-M406分布圖
G-M406在達吉斯坦西南部的Charmalal人中達到最高分布頻率(18%),除此之外在喬治亞(3%)、亞美尼亞(2%)、伊朗(0.5到2.5%)與土耳其(3到6%)等高加索地區更普遍。在黎凡特其被發現有低分布頻率,並遠達埃及與葉門。在歐洲G-M406在希臘最常見(2%),包括克里特島(2.5%),然後在義大利(3%),尤其是西西里島(4%)。G-L293與G-M406都沒有在數百位被檢測的新石器時代歐洲人個體驗出,代表他們自青銅器時代晚期才開始從南高加索擴散。
羅馬人的重新分布
在鐵器時代,G2a人口在印歐語系使用者入侵與其後的塞爾特人戰爭中被淘汰。G2a在尋求避開入侵者進入山中。
古代拉丁人與羅馬人的義大利部落後代在西元前1200年前入侵義大利半島。他們似乎主要屬於單倍群R1b-U152,但攜帶了大量的G2a-L140少數血脈,尤其是G-L13、G-L1264與G-Z1816子分支。拉丁人在義大利中部的家鄉是今天單倍群G2a在歐洲的一處熱點。G2a在拉丁姆的高水平也許是同時由於印歐語系的G-L13,G-L1264與G-Z1816分支的出現與稍早遍布亞平寧山脈並接著被羅馬文明吸收入駐羅馬的新石器時代血脈。
如果來自義大利半島的古羅馬人與其他羅馬化的人群對羅馬帝國其他地區有任何遺傳影響(但他們當然有),那他們一定為帝國疆域內的G2a貢獻了緩和增加(還有R1b-U152與J2)。確實,單倍群G的分布頻率從依著與帝國邊界的距離在減少。單倍群G在今天的北歐與波羅的海國家中比英國更加稀有,儘管事實上農業的傳播同時抵達到這些地區。所以G2a在英國尤其是威爾士的部分(G2a比率最高的地方)毫不意外地會來自羅馬人。另一個理由是德國北部、波蘭與巴爾幹地區的低地森林過於缺乏金屬以至於不曾吸引許多來自高加索地區的青銅器時代工匠。單倍群R1b在東歐北部的百分比率也相對低,更加強了兩個單倍群共同在青銅器時代擴張的假設。
斯基泰人G1
單倍群G1(M342)跟G2b一樣從中東到南亞與中亞都被發現。在新石器時代,一位G2a經過安那托利亞遷移到歐洲,他們的G1表兄弟向東移動到伊朗平原與印度。在歐洲,G1大多限定在羅馬尼亞、摩爾多瓦、烏克蘭西部、波蘭東部、白俄羅斯與立陶宛,一小部分樣本在德國中部與南部。這樣的分布讓人想起R1a-Z93,R1a的印度-伊朗語族分支。青銅器時代與鐵器時代,中亞變成南方G1與J2血脈與北方R1a血脈融合的地方。新的雜交人種就這麼形成,比如一度建立橫跨巴基斯坦到新疆與烏克蘭的帝國的斯基泰人。R1a-Z93、R1b-Z2103、G1、J2b與Q1b等血脈都可能由西徐亞人、薩爾馬提亞人與其他歷史上的草原部落帶入歐洲。
羅馬人也被得知雇傭過斯基泰人或薩爾馬提亞人騎兵進入他們的軍團。根據C. Scott Littleton在他的著作From Scythia to Camelot,幾個圓桌騎士都來自斯基泰人。這個假設也被體現在2004的亞瑟王電影,故事從斯基泰人羅馬騎兵來到不列顛展開。但是,生活在草原的斯基泰人也該帶著夠高的R1a-Z93與R1b-Z2103分布頻率。在不列顛有十幾個家族在英格蘭被確定屬於G1,然後有趣的是一個在威爾士的家族(姓Mills),當地據說是盎格魯薩克遜人入侵後亞瑟王的不列顛羅馬人軍隊撤退的地方。極為稀少的R1b-Z2103在不列顛的多賽特與薩默賽特被發現,包括一位在格拉斯頓伯里(姓Allen,起源於塞爾特,但也可以聯繫到奄蔡人,一個講斯基泰-薩爾馬提亞語言的草原部落),一個與亞瑟王有關的地點。
與單倍群G成對的母系血脈(粒線體單倍群)
與Y染色體單倍群G2一同分化的主要母系血脈是粒腺體單倍群N1a1a、W1與X,都是根源於中東的少數血脈。有趣的是N1a、W(或稱N2b)與X都直接傳承自相當古老的單倍群N*,相當相近巨集單倍群R(HV、JT與UK的祖先,代表歐洲90%的粒線體血脈)。N1a與X的繁衍瓶頸也反映在Y染色體單倍群G2上。這些單倍群被總稱為古歐亞人。
像G2a沒在新石器時代的黎凡特被發現,但主要的G2a人口全部都被發現在安那托利亞與歐洲。40個來自新石器時代安那托利亞的粒腺體樣本被驗出11個人屬於N1a1a,3個屬於X2與1個屬於W1。另外2個屬於甚至今天十分稀有的血脈N1b1a,但與N1a相當關聯。在新石器時代安那托利亞的幾乎一半的粒腺體單倍群血脈與稍微超過一半的Y染色體單倍群血脈因此皆屬古歐亞人。
此外,與現在相比N1a1a與X2同樣被發現在新石器時代歐洲有異常高的分布頻率,N1a1a被發現在線性陶器文化的樣本中佔13%以及X2一般在不同的新石器時代文化中佔5到10%的範圍內。今天,在歐洲人口中發現X2只有1.5%然後N1a1a低於0.5%。現代唯一有粒腺體單倍群X高分布頻率的德魯茲教徒,同樣有超過10%的Y染色體單倍群G。儘管如此,N1a與X無法只與Y染色體單倍群G有所聯繫,因為第一批來自新月沃土的新石器時代農夫都屬於不同父系(與母系)血脈,還包括Y染色體單倍群E1b1b(至少是E-E123)、H2與T1a(或許還有少數的J1與J2)。
 創作內容
創作內容









 未分類 (2)
未分類 (2)